Csirke tojásrakási szindróma, tojásöntési szindróma, EDS-76, "Syndrome-76", csirkék adenovírusos betegsége(Tojáscsepp szindróma - angol) - vírusos betegség, amely a kereskedelmi és tenyésztési tojótyúkokat érinti, a tojástermelés csökkenése, a tojáshéj megpuhulása vagy eltűnése, a héj színének elvesztése a barna tojást tojó tyúkfajtáknál.
Prevalencia. A betegséget először Van Eck írta le 1976-ban. Jelenleg Hollandiában, Angliában, Észak-Írországban, Franciaországban, Olaszországban, Németországban, Spanyolországban, Belgiumban, Jugoszláviában, Dániában, Magyarországon, Svédországban, Argentínában és az USA-ban regisztrálták.
Gazdasági kár a betegség által okozott veszteségek a tojástermelés csökkenésével járó veszteségek miatt. J. Macpherson szerint Angliában a kár évi 2,4 millió fontot tett ki. Francia tudósok megjegyzik, hogy a ketrecben tartott madarak termelékenysége 4-6 héten belül helyreáll, padlón tartva pedig 8-10%-kal alacsonyabb. Az egy madarakra jutó tojásveszteség az érintett állományokban 17,7%, ami a becslések szerint 116,9 penny. A beteg madár legfeljebb 15%-ban torzul, gyenge héjjal vagy depigmentált tojásokkal vagy anélkül. Darbyshire (1980) szerint az érintett peték száma elérheti a 38-40%-ot. J. McFerran és mtsai. (1978) megjegyzik, hogy e betegség következtében a termelékenység csökkenése átlagosan 12 tojás tyúkonként.
A gazdasági károk a keltetőtojások minőségének romlásával is járnak. A tojótyúkból származó több mint 50 tojás nem alkalmas keltetésre a héj törékenysége miatt. A magas selejtezés mellett a csirkék keltethetősége és életképessége is csökkent az élet első napjaiban.
Kórokozó. Különböző kutatók több törzset izoláltak, amelyek azonosságát szerológiai vizsgálatok igazolták. A 127-es törzset Észak-Írországban izolálják a madarak orrüregének és garatának nyálkahártyájából. A vírust (BC 14) vérleukocitákból izolálták, az érintett állományokban lévő csirkék petevezetékéből, ürülékéből, orrgaratából izolált 127-es vírussal való azonosságát szerológiailag igazolták, ezen ágens ellen antitesteket is találtak (W. Baxendale, 1977, 1978; J. McFerran és munkatársai, 1977, 1978). Kísérleti körülmények között a betegséget tojótyúkok és brojlerek megfertőzésével reprodukálták, és megállapították, hogy az izolált vírus etiológiai ágens (W. Baxendale, 1977; W. Baxendale és mtsai, 1978; McCraken, J. McFerran, 1978). ).
A D61 törzset egy tojótyúk kloákájából izolálták Angliában. Magyarországon a 127-es törzzsel azonos B 8/78 törzset kaptunk, Franciaországban 3 törzset izoláltak. A vírust adenovírusként azonosították (Todd, McNulty, 1978), de a fő madáradenovírusokkal rendelkező csoportspecifikus antigéneket géldiffúziós módszerrel nem állapították meg (W. Ba-xendale, 1977; J. McFerran és mtsai, 1978). ). Mindazonáltal az indirekt immunfluoreszcencia módszerével bizonyos fokú keresztreaktivitást figyeltek meg madár adenovírusokkal (W. Baxendale, 1978).
A tojáscsepp vírus nem kapcsolódik a madár adenovírusok 11 szerotípusának egyikéhez sem (J. McFerran et al., 1978), és különbözik a többi madár adenovírustól a csirkék vörösvértest-agglutináló képességében (J. McFerran és mtsai, 1977). , kacsák és libák (W. Baxendale, 1977).
Az ECC-76 vírus magas titerrel szaporodik a kacsavesékben, a kacsaembriómájban és a kacsaembrió fibroblasztjaiban, rosszabb a csirkemájsejtekben és rendkívül alacsonyan a csirkeembrió fibroblasztjaiban. A vírust nem tenyésztik emlőssejtekben. Az adenovírusok első alcsoportjánál leírt módszer szerint replikálódik a sejtmagokban. A fertőzött sejtek vékony metszeteinek elektronmikroszkópos vizsgálata vírusrészecskéket és kapcsolódó zárványtesteket mutatott ki mind az emberi típusú adenovírusokban, mind a madáradenovírusokban.
A kórokozó széles tartományban ellenáll az éternek és a pH-nak. Fertőző tulajdonságai 60°C-on 30 percen belül elvesznek, tripszin, karbamid és piridin is inaktiválja. Ez egy DNS-tartalmú vírus, amelynek nincs külső héja, a cézium-klorid sűrűsége 1,34; a szacharóz ülepedési állandója 940; a virion átmérője 75-80 nm, a kapszid szimmetriája köbös.
V. Kraft et al. (1979) elektronmikroszkóppal tanulmányozták a 127. izolátumot, a CsCl gradiensben három vonalat figyeltek meg. Két vonal sűrűsége 1,32 g/ml volt, amelyeket nem választottunk el frakcionálással, és amelyek felelősek a magas hemagglutinációs aktivitásért és fertőzőképességért. Ezek a vonalak az adenovírusokra morfológiailag jellemző részecskéket tartalmaztak. A harmadik vonal (sűrűsége 1,30 g/ml) főleg törött részecskékből állt, és a hemagglutinációs aktivitás észrevehető csúcsát mutatta.
A vírus hemagglutininje 70°C-on 30 percig stabil, de magasabb hőmérséklet gátolja az aktivitást. A tisztított oldható hemagglutinin 2 db 65 000 és 67 000 molekulatömegű polipeptidet tartalmaz, 25-30 nm hosszúságú rúd alakú struktúrákból áll.
A vírus ikozaéderes, az adenovírusokra jellemző a részecskeméret, a kapszid szerkezet, a kapszomerek száma (V. Kraft et al., 1979).
epidemiológiai adatok nem tanult eleget. A tojáscsepp szindróma minden fajtájú tojótyúkot érint 25 és 35 hetes kor között. A legfogékonyabbak a barna tojást termelő húsfajták, valamint a tenyészmadarak. A fehér tojást tojó fajták kevésbé érzékenyek. A tojásleadás szindróma vírusáról azt gondolják, hogy kacsa eredetű, de nem patogén a kacsákra. Az észak-írországi csirkék feltehetően azután fertőződtek meg, hogy beoltották őket Marek-kór ellen a hollandiai kacsaszövettenyészetben előállított Rispens törzsből. Írországban és Németországban figyelték meg először az ECD-76 jeleit mutató betegséget Hollandiából importált csirkékben.
A betegség a madár kötőhártya és szájon át történő mesterséges fertőzésével szaporodik. A vírus függőlegesen terjed a tojáson keresztül. Számos kutató felismeri a fertőzés átvitelének horizontális útvonalát is, nagy jelentőséget tulajdonítva az alomnak. A vírus terjedése a kakasok spermájával várható.
A tojáscsepp szindróma epizootológiai jellemzői- a vírus azon képessége, hogy nem nyilvánul meg a madarak szervezetében, amíg el nem érik a pubertást. A kórokozó aktiválódásának oka a szervezetet megterhelő hatás a peterakás megindulása miatt.
Patogenezis nem kellően tanulmányozott. Csak az ismeretes, hogy megsértették a képződési mechanizmust tojáshéj. Petefészek sorvadás és vérzés is megfigyelhető. A méhben megnövekszik a savasság, ami a kalcium feloldódásához és a héjképződés megzavarásához vezethet. A tojótyúkok kísérleti fertőzése során a 4-7. napon hibás tojások jelennek meg, a vér fehérje- és kalciumtartalma nem változik.
A klinikai tünetek általában nem jelennek meg. Előfordulhat azonban felborzolt toll, vérszegénység, hasmenés, elesett állapot, a tojásrakás során csökkent aktivitás. A betegség későbbi stádiumában a madarak 10-70%-ánál jelentkezhet cián- és fésűs cianózis. A betegség jellemző jelei a tojások héjának elszíneződése barnára, vékony héjú vagy anélküli tojás megjelenése, a tojástermelés csökkenése. Ezek a tünetek 2-3 héttől 6-12 hétig fennállnak, és a csirkékre jellemzőek. húsfajtákés barna tornacipőt. A tojásfajtákra jellemző a tojás belső minőségének változása is: a fehérje vizenyőssé, zavarossá válik.
Patológiás elváltozások vagy hiányzik, vagy gyenge hurutos bélgyulladás, sorvadás, ödéma és a méh mirigyszövetének sejtjei és a petevezetékek mononukleáris infiltrációja van.
Diagnózis és differenciáldiagnózis. A diagnózis ezeken a klinikai tüneteken, kóros elváltozásokon és szerológiai vizsgálaton alapul. A legmegbízhatóbb diagnosztikai módszer a hemagglutinációs gátlási teszt (HITA). A párosított szérumok vizsgálatakor 15 nappal és 2-3 héttel a fertőzés után az antihemagglutininek titerének 1: 1280-1: 2560-ra történő növekedését figyelték meg.
A tojásleadás szindrómát meg kell különböztetni a Newcastle-betegségtől, a fertőző hörghuruttól, kokcidiózistól, helmintiázistól, a peszticidekkel, gombaölő szerekkel, mikotoxinokkal való mérgezéstől és a tojástermelés csökkenését okozó különféle nem fertőző etiológiájú tényezőktől (tartási körülmények, takarmányminőség, kiegyensúlyozott étrend, stb.).
Kezelés nem fejlődött. Az utódok kikelhetőségének és életképességének optimális szintjének megőrzése érdekében teljes értékű étrend alkalmazása javasolt, különösen az esszenciális aminosavak, vitaminok és nyomelemek tekintetében.
Immunitás és a specifikus profilaxis eszközei. A természetesen beteg madár erős immunitást fejleszt ki. A fertőzés után 7 nappal kimutathatóak az IgG immunglobulinhoz kapcsolódó semlegesítő, antihemagglutináló és kicsapó antitestek. A fertőzött tyúkok tojásából kikelt csibék megtartják az anyai antitesteket, amelyek felezési ideje 3 nap. A passzív antitestek biztosítják a csirkék fertőzésekkel szembeni rezisztenciáját az élet első 4 hetében.
Specifikus profilaxisként a kacsaembrionális sejttenyészetben tenyésztett BC 14 törzsből olajalapú formol vakcinát használnak. A vakcinát egyszer, intramuszkulárisan vagy szubkután adják be 0,5 ml-es (1000 HAU) dózisban, 18 hetes kortól kezdve.
Az SSJ-76 elleni vakcinát Franciaországban, Hollandiában, Nagy-Britanniában, Belgiumban és Luxemburgban, Argentínában tesztelték. A vakcinázást követően a virémiás fázis és az ezzel járó víruskiválasztás nem figyelhető meg, a tojástermelés és a héj minősége javul. Az immunitás 15 nap után alakul ki, a maximális antihemagglutinin 30 nap után halmozódik fel. Az antitest-titer 12-16 héttel az oltás után csökkenni kezd. 40-50 hét után az antitestek nem mutathatók ki.
A vakcinázást átmeneti intézkedésnek tekintik a vírus terjedésének és a csirkékhez való alkalmazkodásának veszélye miatt. Jugoszláviában (M. Krecov et al., 1980) a 127-es vírustörzsből készült, 13-propiolaktonnal inaktivált és alumínium-hidroxiddal adszorbeált vakcinát pozitív eredménnyel tesztelték.
Megelőző és ellenőrző intézkedések. A keltetéshez 40 hetesnél idősebb tyúkok tojásait ajánljuk, amelyek negatív eredményt adnak a szerológiai vizsgálatokban. Angliában, Észak-Írországban és Németországban a szerológiai reakciókban pozitívan reagáló madár levágását radikális intézkedésnek tekintik a tojáscsepp szindróma leküzdésében és megelőzésében annak érdekében, hogy megakadályozzák a kórokozó függőleges átvitelét a tojáson keresztül. Inaktivált vakcina használata megengedett.
Az adenovírusfertőzések különféle madarak (csirke, pulyka, gyöngytyúk, fácán, pézsmakacsa, galamb, fürj, törpepapagáj) betegségei. A madár adenovírusok 12 típusa okozza, amelyek nem rendelkeznek közös antigéncsoporttal más humán, majom, sertés és egér adenovírusokkal.
A madár adenovírusok az adenovírusok családjába tartoznak, mivel számos közös tulajdonsággal rendelkeznek ezzel a családdal: részecskeméret, virion szerkezet, genom szerkezet, replikációs típus. Az adenovírusokban a virion mentes a külső lipoprotein héjtól, nem tartalmaz lipideket és glikoproteineket, gömb alakú kapszid alakú, mérete 70-80 nm.
A madarak adenovírusos fertőzései a következők: СELО - fertőzés, EJS-76 - tojáscsepp szindróma, adenovírus légúti betegség, hepatitis zárványtestekkel.
Tojáscsepp szindróma- a tojótyúkok betegsége, amelyet a tojáshéj felpuhulása, hiánya (leöntése) vagy depigmentációja jellemez. A tojástermelés jelentős csökkenése kíséri.
A betegséget először Hollandiában írták le 1976-ban J. Van Ecke és munkatársai. Az elvégzett vizsgálatok lehetővé tették a madarak NB, ILT kizárását. Előtte a világon senki sem figyelt a tojástermelékenység csökkenésének jellegzetes dinamikájára. Az 1976 előtt csirkékből származó vérszérum egyik múzeumi mintájában sem találtak EDS-76 vírus elleni antitesteket. Ez vitathatatlan megerősítése annak, hogy a betegség kialakulása egy bizonyos időszakra korlátozódik.
Számos országban különböző időpontokban izolálták az EDS-76 vírus törzseit: BC-14 az Egyesült Királyságban, B / 78 Magyarországon, E-77 Olaszországban, 3877 Franciaországban, J RA-1 Japánban. Az összes kapott izolátum szerológiailag teljesen azonos volt az első izolált -127 törzzsel.
Az EDS-76 elterjedt a fejlett ipari baromfitechnológiával rendelkező országokban, és nagyon jelentős gazdasági károkat okoz, különösen a tenyésztelepeken. A veszteség a tojótyúkok tojócsúcs idején elveszett tojásaiból áll (egyes adatok szerint - tyúkonként akár 15 tojás, mások szerint - akár 25%, az USA-ban - a kár elérte az 5 millió dollárt), valamint a kereskedelmi tojások kereskedelmi minőségének csökkenéséből és a tenyésztési rétegekből (fejenként legfeljebb 50 tojás volt a selejtezés). A betegeknél a tojástermelés általában nem áll vissza az eredeti szintre. Jelenleg a betegséget kellőképpen tanulmányozták, azonban az etiológia, az epizootológia és a patogenezis kérdései nagyrészt ellentmondásosak.
Kórokozó- Az adenovírusok családjába tartozó DNS-tartalmú vírus. 1976-ban izolálták Észak-Írországban beteg madarak orrüregéből és petefészkéből, jelenleg 127-ként ismerték, és az EDS 76 kórokozójaként azonosították.
Számos tudós véleménye szerint az EDS-76 vírus egy kacsa adenovírus, amely nem okoz patológiát a természetes "gazdaszervezetekben". A betegség először Hollandiában, az intenzív kacsatenyésztés országában jelent meg. A csirkék elsődleges fertőzését ezzel mesterségesen állították elő a Rispens törzsből származó Marek-kór elleni vakcina bejuttatásával, amelyet adenovírussal fertőzött kacsaembrionális fibroblaszt sejttenyészet felhasználásával állítottak elő. A vírus gyorsan alkalmazkodott az új "gazdaszervezethez" - a csirkékhez, és megmutatta patogén tulajdonságait.
Az ECC-76 kórokozó tulajdonságai teljes mértékben megfelelnek más madár adenovírusok jellemzőinek, kivéve a csirkék, kacsák és libák vörösvértest-agglutináló képességét. Ezen az alapon ez még inkább "madár adenovírus", mint más adenovírusok, amelyek nem rendelkeznek hemagglutináló képességgel.
A vírus instabil a hőre: +65 C hőmérséklet 30 perc alatt teljesen inaktiválja a vírust. Az EDS vírus 15-ször érzékenyebb az UV sugárzásra, mint a fertőző bronchitis vírus. Az ECC-76 vírus ellenáll a táptalaj pH-értékének változásainak, és különösen ellenálló lúgos környezetben. Ellenáll az ismételt fagyasztásnak és felolvasztásnak, más fizikai tényezők hatásainak, valamint fertőtlenítőszereknek.
Epizootológia. Az EDS-76 leggyakrabban minden fajtájú tojótyúkot érint, a betegség maximális manifesztációjával a 26-35 hetes korban tapasztalható megnövekedett peteérés időszakában. Különösen fogékony baromfihúsfajták.
Megállapították, hogy a 40 hetesnél idősebb madarak nem szenvednek EDS-76-tól, és nem választják ki a vírust, de a vér e vírus elleni antitesteket tartalmaz.
Az adenovírusok széles körben elterjedtek a házi- és vadkacsák körében is, amelyek ugyanakkor nem is betegszenek meg. Magyarországon a kacsák 85%-ában és a libákban 65-100%-ban mutattak ki ellenanyagot. Természetes és vadon élő körülmények között a természetben a kórokozó tározói. A betegség csekély fertőzőképessége ellenére fennáll annak a veszélye, hogy a kórokozót vadon élő madarak: galambok, verebek juttatják be a gazdaság területére.
Az EDS-76 kórokozó fő forrása a beteg csirkék, amelyek széklettel választják ki a vírust, vagy tojással továbbítják azt utódjaiknak.
Az adenovírus terjedésének fő útvonala függőleges. Kisebb jelentőségű a kórokozó átvitelének horizontális és kontaktus útja. A betegség gyorsabban terjed, ha a csirkéket a padlón tartják.
A betegség járványtani jellemzője a kórokozó újraaktiválódásának szigorúan meghatározott ideje, miután a fertőzött madár eléri a teljes ivarérettséget. Ennek oka lehet a stressz, amely a tojótyúkok szervezetének élettani átrendeződése a peterakás aktív szakaszának kezdete előtt. A betegség kialakulásának egyéb specifikus hajlamosító tényezőit az életkor kivételével nem állapították meg.
Patogenezis. A látens fertőzés időszakában a vírus megmarad a bélben. A vírus újraaktiválódása a peterakás kezdetén bekövetkezett hormonális profilváltozás miatt következik be, amelyet stressztényezőnek tekintenek. Nyilvánvalóan a vírus közvetlen hatással van a méh mirigyhámjára, ami héj nélküli vagy vékony héjú tojások lerakásához vezet.
Klinikai tünetek. Az ESS-76-nak nincsenek jellegzetes tünetei. A madár étvágya csökkent, tollazata felborzolt, hasmenése van. A madár depigmentált, deformálódott és hibás tojásokat hordoz 2-3 hétig. Jelentősen növeli a "márvány" tojások számát, növeli a harc és a bevágások százalékos arányát. A tojásfehérje vizes és zavaros.
A tojás termelékenysége átlagosan 15-30% -kal csökken, egyes esetekben akár 50% -kal. A termelékenység csökkenése 5-6 hét alatt fokozatosan növekszik, ezt követően a csirkék sejttartalmával nagyon lassan helyreáll a produktivitás, padlóval nem.
Egy madár elpusztulása az EDS-76 során, még a betegség csúcspontján is, jelentéktelen. A tojótyúkok mortalitása eléri a 3-5%-ot, főként a tojássárgája hashártyagyulladása, csípés miatt.
Ha az embriókat vírussal fertőzték meg, elpusztulásuk a fertőzést követő 4-6. napon kezdődik, és a fiókák kikeléséig tart.
Patológiás elváltozások elsősorban a reproduktív traktus szerveiben lokalizálódnak, és petefészek-sorvadás formájában fejeződnek ki, néha vérzések találhatók bennük. Minden esetben kisszámú érlelő és érett tüsző található a petefészekben. A petevezeték általában lerövidül, fala elvékonyodik, hiperémia gócokkal. A máj megnagyobbodott, ödémás, petyhüdt állagú lehet.
Diagnosztika. Az előzetes diagnózist a baromfiipar járványtani felmérésének adatainak átfogó értékelése alapján állítják fel. Ugyanakkor kiemelt figyelmet fordítanak a kifejlett tyúkok tojástermelésének dinamikájára, a naposcsibék tömeges pusztulására. Fontos diagnosztikai érték a madarak életkora a tojástermelés visszaesése során.
A petevezeték érintett területeinek szövettani vizsgálata intranukleáris testeket - hámsejtek zárványait - tárja fel.
A laboratóriumi diagnosztika a vírus kóros anyagból történő izolálásán (lehetőleg kacsaembriókon) és az RZGA-ban, MFA-ban történő azonosításán alapul.
A szerológiai diagnosztikát (RZGA, RDP, ELISA) használják az EDS-76 vírus elleni antitestek dinamikájának kimutatására madarak páros vérszérumában, valamint az immunitás intenzitásának megállapítására.
Az EDS-76 differenciáldiagnózisában a tojástermelés CELO-vírusok, fertőző hörghurut, Newcastle-betegség vagy a fogvatartási feltételek megsértése által okozott csökkenése kizárt.
Megelőző és ellenőrző intézkedések. Az EDS szempontjából kedvezőtlen ponton az állat-egészségügyi intézkedések szigorú betartása kötelező a kórokozó fertőzés forrásból való terjedésének kizárása érdekében. Fertőtlenítse a befúvó és elszívó szellőzőrendszert.
Kedvező prognózis csak speciális profilaxis alkalmazásával lehetséges. A madarakat 100-110 napos korukban egyszer intramuszkulárisan folyékony, inaktivált adszorbeált vakcinával immunizálják ECC-76 ellen. Ásványi és olajadjuváns alapú (fertőző bronchitis, IBD, Newcastle-kór és EDS-76 elleni) vakcinák monovalens és kapcsolódó formáit fejlesztették ki és vezették be a baromfitenyésztés gyakorlatába.
Tojáscsepp szindróma-76 (ESD-76)- vírusos betegség, amelyet a csirkék reproduktív rendszerének károsodása, a tojástermelékenység csökkenése és a tojás minőségének romlása jellemez.
Az EDS-76 okozta kár a tojótyúkonkénti átlagosan 10-30 tojás hiányából adódik (tenyésztyúkokból átlagosan 50 tojást selejteznek), héj nélküli tojások, vérgyűrűs tojások, a csirkék keltethetőségének csökkenése és alacsony életképességük két hetes korig.
Etiológia. A betegség kórokozója az Aviadenovirus nemzetségbe, az Adenoviriclae családba tartozó vírus, amelyet bizonyos fizikai-kémiai és biológiai tulajdonságok alapján kacsa adenovírusként azonosítottak. A virion köbös szimmetriájú, nincs szuperkapszid héja, átmérője 80 nm, az arcon 6 kapszomer található. Az ikozaéder csúcsain gomba alakú megvastagodások észlelhetők. Úgy gondolják, hogy a vírus felületi antigénjének - hemagglutinin - hordozói. A vírus genomja DNS-ből áll. A vírus molekulatömege 170-175 megadalton, ülepedési állandója 560, a cézium-klorid gradiensben lebegő sűrűsége 1,32-1,35 g/cm 3 . A vírus ellenáll az éternek, a kloroformnak és a hőmérsékleti tényezőknek. Ha a vírust tartalmazó anyagot 30 percig 56 °C-ra melegítjük, a fertőző aktivitás legfeljebb 50%-kal csökken. A 70°C-ra való melegítés 10 perc elteltével a fertőző és hemagglutináló aktivitás elvesztését okozza. Fagyott állapotban -25°C-on a vírus több mint 3 évig életképes marad. A vírus viszonylag stabil pH 6,0-9,0 között. 50%-os glicerinben -30 -50°C hőmérsékleten a kóros anyagban lévő vírus több évig aktív marad. A vírus jól tárolható liofilizált állapotban. A madarak testében a vírus lokalizációjának vezető helye a reproduktív rendszer szervei, de a látens fertőzés időszakában a belekben és más szervekben is fennmaradhat. A vírus a petevezetékből, leukocitákból izolálható
beteg csirkék vére, felső légutak, orr- és garatnyálka, máj, kloáka tartalma a betegség első klinikai tüneteinek megjelenésétől számított 3-5 napon belül. A kísérletileg fertőzött madarak belső szerveiben és szöveteiben a vírus legfeljebb 5 hétig fennmarad. A székletben a fertőzést követő 2 héten belül kimutatható. Az EDS-76 vírus antigén változatait és típusait nem állapították meg. Beteg csirkékben a fertőzést követő 6-8. napon szérum antitestek képződnek, amelyek vírussemlegesítő, antihemagglutináló és kicsapó tulajdonságokkal rendelkeznek. Az antitesttermelés dinamikájában két csúcs van. Az első (JgM-csúcs) a fertőzést követő 10-11. napon, a második (JgG-csúcs) a fertőzést követő 16-18. napon észlelhető. A semlegesítő antitestek a fertőzést követő hatodik napon kezdenek képződni, a titer 28 napig emelkedik. A fertőzést követő 6. naptól kicsapódó antitestek is képződnek, de a 17. naptól titerük csökkenni kezd. A szeropozitív szülőktől származó csibék 4 hetes korukig 3 napos felezési idejű anyai antitesteket tartalmaznak. De nem tudják folyamatosan megvédeni a madarat az adenovírusoktól, amelyek függőlegesen továbbítják az utódokat, és stresszes helyzetekben kezdenek megnyilvánulni, beleértve a peteérés csúcsának elérését. Az ECC-76 vírus nem tartozik a jelenleg ismert madáradenovírusok 12 szerotípusa egyikébe sem, és velük ellentétben képes csirkék, kacsák, pulykák és libák vörösvértesteinek agglutinációjára.
járványtan. A tojótyúkok minden fajtája fogékonyak a betegségre, a betegség maximális manifesztációjával a maximális tojástermelés időszakában (180-140 nap), de a betegség a termelési ciklus bármely szakaszában előfordulhat. A nagyon produktív hús-tojás és tojás keresztezett csirkék érzékenyebbek a betegségre. Az EDS-76 vírus széles körben elterjedt a házi- és vadkacsák, valamint a libák körében, számukra apatogén, és nem okozza a betegség klinikai megnyilvánulását. A kórokozó perzisztenciáját általában visszamenőleg igazolják specifikus antitestek jelenléte, de sok esetben kacsából és libából is izolálható a vírus. A kislibák EDS-76 vírussal való kísérleti fertőzését követően a kórokozó 3 héten belül izolálható a fertőzött madarak ürülékéből, ami indokolja, hogy a libákat potenciális fertőzési forrásként tekintsük. Vadludak, gémek, verebek, hattyúk, baglyok, gólyák vérszérumában kimutathatók az EDS-76 vírus elleni antitestek, de a betegség klinikai megnyilvánulásáról ezen madárfajok körében nincs információ. A kórokozó terjedésének vezető útja a transzovariális. A tojások különösen intenzíven fertőződnek a virémia időszakában, amely időben egybeesik a tojástermelés csökkenésével. Nem kizárt a kórokozó átvitele a kakas spermájával. A kórokozó vízszintes terjedése a betegség kezdeti szakaszában figyelhető meg, és akkor a legkifejezettebb, amikor a baromfit a padlón tartják. Ebben az időszakban a vírus intenzíven ürül a széklettel, garat- és légcsőnyálkával, szennyezi a takarmányt, a környezetet, ami a fertőzött takarmány és ürülék elfogyasztásával hozzájárul az állomány újrafertőződéséhez. A betegség járványtani jellemzője egyes keresztezett csirkéknél a fertőzés egyenlőtlen megnyilvánulása egy gazdaság különböző baromfiházaiban, ami az EDS-76 szerint kedvezőtlen. Ezenkívül előfordultak olyan esetek, amikor ugyanabban a baromfiházban az érintett madarakkal együtt jelentős számú tojótyúk maradt, fenntartva a normál tojástermelést a teljes termelési időszak alatt.
Klinikai tünetek. A betegségnek nincsenek jellegzetes tünetei. Hasmenés, némi elnyomás, csökkentett takarmányfelvétel lehetséges. A betegség csúcspontján - gyengült légzés, folyékony zöldes alom, 1-2 héten belül megfigyelhető. A betegség későbbi szakaszában a fülbevaló és a fésű cianózisa lép fel. Az EDS-76-ban szenvedő madarak mortalitása még a betegség akut formájának csúcspontján is jelentéktelen.
Ritka esetekben a felnőtt rétegek között elérheti a 3-5%-ot, főként a sárgája hashártyagyulladása és kloacitise miatt. A betegség vezető tünete a tojástermelékenység 15-30%-os csökkenése, egyes állományokban pedig 50%-kal vagy még ennél is nagyobb mértékben. A tojástermelés csökkenése bizonyos mintázat szerint megy végbe: a teljes csökkenési időszak 6-7 hét, ebből 4-5 hét közvetlenül a csökkenésre, 2-3 hét pedig a gyógyulási időszakra esik. Ketrectartalommal a csirkék termőképessége szinte teljesen helyreáll, vagy 1-3%-kal alacsonyabb marad a normánál, padlótartalom mellett pedig általában 7-10%-kal kevesebb, mint a csökkenést megelőző tojástermelés. Az EDS-76-ból való felépülés során a madár 6-8 hétig héj nélküli („tojásöntés”) vagy depigmentált, vékony héjú tojásokat visz magával, gyűrűs vagy csíkos, durva képződményekkel a felszínen. Jelentősen megnő a "márvány" tojások száma, nő a harci és rovátkolt tojások aránya. A tojások kotlási tulajdonságai romlanak, míg termékenységük, keltethetőségük és a csirkék életképessége az élet első két hetében csökken. A héj és az ún. "zsíros tojás" minőségének változása leggyakrabban színes keresztezések rétegeiben és brojlercsirkékben fordul elő. A fehér keresztes tyúkoknál a tojás fehérje része elsősorban helyi cseppfolyósodás formájában változik.
Patomorfológia. A madarak belső szerveinek kóros és anatómiai változásai az EDS-76-ban rendkívül gyengén fejeződnek ki. Akut formában bizonyos esetekben atrófiás változásokat észlelnek a petefészekben, a tüszők számának és méretének csökkenését, hiperémiát vagy vérzést a tüszők kötőszöveti tokjában, deformációjukat és degenerációjukat. A petefészekben vannak olyan területek, ahol a tüszők teljesen elfajultak, és sárgásbarna színű nekrotikus tömeggé alakultak. Talán a ciszták megjelenése. A petevezeték általában rövidebb és vékonyabb, mint az egészséges madaraké. A nyálkahártya hiperémiája és petechiális vérzése, duzzanata feltárul, legkifejezettebb a petevezeték méhrészében, ritkábban a hüvelyi és fehérjerészekben. Egyes esetekben a tojássárgája hashártyagyulladása figyelhető meg. A máj néha megnagyobbodott, petyhüdt szerkezetű és sárgás színű. Ugyanakkor az epehólyag növekedése, túlcsordulása vizes epével, világoszöld színű. A madarak limfoid szerveinek az EDS-76 vírus általi legyőzésével kapcsolatos információk ellentmondásosak.
Diagnosztika. A diagnózis felállítása epizootológiai, klinikai, patoanatómiai adatok és laboratóriumi eredmények alapján történik, amelyek magukban foglalják: a vírus izolálását kacsaembriókon, kacsaembrió fibroblaszt sejttenyésztését vagy kacsa- vagy csirkeembriók májszövetének tenyésztését, majd az embriók azonosítását. a kórokozó az RHA, RHA, MFA, ELISA, csirkéken végzett biológiai vizsgálatban; specifikus antitestek meghatározása tojótyúkok vérszérumában vagy tojássárgájában RZGA vagy ELISA módszerrel; a reproduktív traktus szerveinek szövettani vizsgálata.
A virológiai vizsgálathoz petefészek mintákat tüszővel, petevezetéket, végbélt tartalommal, kloáka tampont, véralvadásgátlóval (heparinnal vagy 1%-os nátrium-citrát oldattal) tartalmazó vért használnak. A kóros anyag mintavételét a betegség kezdetét követő első 3-5 napon belül, de legkésőbb a madár klinikai halálát vagy levágását követő 2 órában kell elvégezni. Az 1-1,5 cm-es szervdarabokat kémcsőbe vagy fiolába helyezzük 5,0-8,0 ml sóoldattal vagy Hank-oldattal, amely 1 ml-enként 1000 NE penicillint, 1000 μg sztreptomicint és 50 nystatint tartalmaz. Stabilizátorként 0,5-1% szarvasmarha-szérumalbumint vagy 0,5% zselatint célszerű hozzáadni az oldathoz, hogy megakadályozzuk a vírus inaktivációját. A köpenytörlőket steril vattacsomóval veszik, és a fenti adalékokat tartalmazó sóoldatba merítik. A vért általában a brachialis (axilláris) vénából, a jugularis vénából vagy szúrással veszik a szívből. A további vizsgálatokhoz a leukocita frakciót teljes vérből izolálják. Ugyanakkor minden madártól etetés előtt 10-15 ml vért veszünk, néhány csepp heparint vagy nátrium-citrátot adunk hozzá, és 1000 fordulat / perc sebességgel centrifugáljuk. 30 percig, majd a buffy coat-ot pipettával leszívjuk. A kapott leukocitákat steril sóoldatba visszük át, ahol 1 ml oldathoz 100 NE penicillint, 100 µg sztreptomicint adunk, és több ciklusos fagyasztásnak és felengedésnek vetjük alá a sejtek elpusztítása érdekében. A vírus izolálására romlott leukociták szuszpenzióját használják. A vírusizolálási eljárás megkezdése előtt a kóros anyagot meg kell szabadítani a tartósítószertől, amit a minták ismételt, 0,15 mol/l nátrium-klorid tartalmú, 7,0-7,2 pH-értékű, steril sóoldattal történő ismételt mosásával érnek el. Ezután a petevezeték vagy béldarabokat a tartalommal együtt összetörjük és steril kvarcüveges mozsárban vagy homogenizátorban őröljük. A zúzott anyagból 10%-os foszfát pufferoldatban készült szuszpenziót készítünk, és 1500-2000 fordulat/perc sebességgel centrifugáljuk. 30 percen belül. A felülúszót pipettával leszívjuk, steril fiolákba töltjük, és 1 ml-enként 200 egység penicillint, 100 μg streptomycint és 20 egység nystatint adunk hozzá. Az antibiotikumokkal való érintkezés szobahőmérsékleten 45-60 perc. A kapott anyag sterilitásának ellenőrzése érdekében az oltást BCH, MPA, Kitt-Tarozzi táptalajra és Sabouraud táptalajra végezzük, majd a növényeket 10 napig inkubáljuk 37°C-on (és a Sabouraud táptalajt 20-22°C hőmérsékleten).
Szerológiai vizsgálatokhoz vérszérumot küldenek 1-120, 160-180, 220-140, 300 napos madarakból, valamint olyan madarakból, amelyek életkoruk szerint nem használhatók, valamint a betegség klinikai tüneteit mutató csirkékből (20- 15 minta minden csoportból). A 3,0-4,0 ml térfogatú vérmintákat külön kémcsövekbe gyűjtjük, steril sóoldattal előzetesen megnedvesítjük, majd 20-30 percre 37°C-os termosztátba helyezzük. A keletkezett alvadékot fémtűvel vagy üvegrúddal felkarikázzuk, és hűtőszekrényben 4-6°C-on legalább 3-4 órán át tároljuk. A leülepedett szérumot tiszta számozott kémcsövekbe öntik, és gumidugóval lezárják. Munkavégzés előtt a szérumot 56°C-os vízfürdőben 30 percig melegítjük. a termolabilis inhibitorok inaktiválására.
Gyanús madaraktól származó tojások kutatási célú küldésekor célszerű a nem megfelelő (vízkőtelenített, depigmentált) tojásokat kiválasztani. A kivonatok elkészítéséhez vegyünk 1,5 ml sárgáját, és szuszpendáljuk egy kémcsőben 6,0 ml 7,2-7,4 pH-jú sóoldattal. Ezután a gumidugóval lezárt kémcsövekbe 2,0 ml diklór-etilént (kloroform) és 1,0 ml étert adunk, az elegyet rázással intenzíven keverjük, 1 órán át 37 °C-on melegítjük, alkalmanként megrázzuk és centrifugáljuk. 15 percig. 2000 ford./percnél. A kémcsövekben kapott átlátszó, enyhén opálos felülúszó a kezdeti 1:5 hígítású sárgája kivonat, amelyet vizsgálati anyagként használunk.
A vírus izolálásához 10-12 napos inkubációjú kacsaembriókat használnak, amelyek nem tartalmaznak EDS-76 vírus elleni antitesteket. Fertőzés előtt az embriókat ovoszkóposan átvizsgálják, munkavégzésre alkalmas, mozgékony, fejlett érrendszerrel rendelkező embriókat választanak ki. A fertőtlenítés után a héjban történő fertőzéshez két lyukat ütünk ki: az egyiket a pugi közepén, a másikat a tojás oldalsó felületén a pugi határa alatt 2-3 mm-rel az előre megjelölt vaszkuláris térben. A vizsgálati anyag 0,2 ml térfogatú elkészített szuszpenzióját fecskendezzük be minden embrióba az oldalsó nyíláson keresztül az allantois üregbe. Ugyanakkor az anyag minden egyes mintájával legalább 4-5 embriót fertőznek meg. Fertőzés után a héjon lévő lyukat paraffinnal töltik ki - először az oldalát, majd a tetejét. A fertőzött és a kontroll embriókat 37 °C-on 96-120 órán keresztül inkubáljuk. Az inkubáció során az embriókat 24 óra elteltével naponta egyszer ovoszkópos vizsgálatnak vetik alá. A későbbi időszakokban elpusztult és a fertőzést követő hatodik napon életben maradt embriókat 13-18 órán át 4-6°C-ra hűtjük, majd kinyitjuk. Minden felnyitott embrióból extraembrionális folyadékot gyűjtünk, 1 ml-enként 200 NE penicillint, 200 µg streptomycint és 20 NE nystatint adunk az antibiotikumokhoz, majd az érintkezés után a tesztanyag második passzálását végezzük. azonos sorrendben fejlődő embriókról. Ezután hajtsa végre a harmadik szakaszt. A harmadik passzázs embrióiból gyűjtött extraembrionális folyadékot RGA-ban és RZGA-ban vizsgálják a hemagglutináló ECC-76 vírus indikációja és azonosítása céljából. A vírusos hemagglutinin intenzív felhalmozódása az embriók szerveiben és szöveteiben a fertőzés után 2-3 nappal kezdődik. 5-6 napra a vírustiter eléri a maximális értéket, esetenként 15-16 log HAU/02 ml-t. Ebben az esetben az embriók elpusztulása a vírus hatására meglehetősen ritkán (1-2%) fordul elő. A vírus fejlődő csirkeembriókon tenyészthető, de felhalmozódása kevésbé intenzív, függetlenül az adaptív passzázsok számától. Ezenkívül a vírus szaporodását a fertőzött kacsaembriókban az egymást követő passzálások során a szervekben és szövetekben kialakult kóros elváltozások természete szabályozza. Az embriók növekedése és fejlődése elmarad, a fej és a nyak bőrének hiperémiája és duzzanata, petechiális vérzések a bőrön és a bőr alatti szövetekben, a CAO elhomályosodása, a máj és a vesék enyhe növekedése.
A vírust különféle sejtkultúrákon és madarak szerveinek és szöveteinek explantátumainak szuszpenzióin is tenyésztik. Gyakrabban kacsaembriók fibroblasztjait és csirkeembriók máj- és vesesejtek tenyészeteit használják. Elsősorban tripszinezett kacsafibroblasztok 24-48 órás tenyészetét használjuk, amelyet hagyományos módszerrel állítanak elő és üvegfiolákban (matracokban) vagy kémcsövekben tenyésztenek. A vizsgálati anyagot fiziológiás sóoldattal 10-1-re és 10-3-ra hígítjuk, majd 3 egyrétegű fiolát fertőzünk meg az anyag minden egyes hígításával 3 ml-es térfogatban. A fertőzött és kontroll fiolákat 37-38°C-on inkubáljuk. A citopátiás hatás figyelembevétele érdekében az injekciós üvegeket kis nagyítású mikroszkóp alatt 24 óránként 7 napon keresztül nézzük.
Az első citopátiás elváltozások a tenyészetek fertőzése után 36-48 órával jelentkeznek, és a sejtmembránok megvastagodása, maguk a sejtek megnagyobbodása és lekerekítése jellemzi őket. A fertőzés után 96 órával az egyrétegű réteg elpusztul, sok elhalt sejt lebeg a táptalajban, míg egy részük hosszú folyamatokkal kapcsolódik össze. Ha a tenyészeteket hematoxilinnel és eozinnal festjük, nagy szemcsés bazofil intranukleáris zárványok láthatók bennük, amelyek összeolvadnak a bazofil tömeg formálatlan felhalmozódásaival. Nagyszámú óriás többmagvú sejt található, amelyek 8-10 részben elpusztult sejtmagot tartalmaznak. Ez utóbbiak vakuolákat képeznek a nukleáris anyag maradványaival. Az egyes sejtek citoplazmájában vakuolák képződése és eozinofil granularitás figyelhető meg. Az ECC-76 vírus felhalmozódik a tenyésztőfolyadékban, ezért a benne lévő vírusrészecskék magasabb koncentrációjának elérése érdekében a fertőzött sejttenyészeteket a vírus felhalmozódásának végső szakaszában három egymást követő fagyasztásnak és felengedésnek vetik alá, a kapott szuszpenzió felhasználásával további vizsgálatok a hemagglutináló ECC-76 vírus kimutatásakor.
A vírus indikációjára és azonosítására leggyakrabban RHA-t, RZGA-t, valamint MFA-t, ELISA-t és PCR-t alkalmaznak.
A fejlődő embriókban vagy sejttenyészetekben a vírus kimutatásának vezető módszere a vírusos hemagglutinin kimutatása az RHA-ban csirke eritrociták 1%-os szuszpenziójával az extraembrionális vagy tenyészfolyadékban. Kezdetben hozzávetőleges RGA-t helyezünk üvegre vagy Petri-csészékre 1%-os eritrocita-szuszpenzióval sóoldatban. Egy csepp eritrocita szuszpenziót és egy csepp vírustartalmú anyagot viszünk fel egy jól megmosott és zsírtalanított üvegfelületre, összekeverjük, és megfigyeljük agglutinált eritrociták pelyheinek megjelenését. A pozitív RGA 3-5 percen belül megjelenik, és agglutinált vörösvértestek pelyhek képződése jellemzi. A pozitív RGA eredményeit az előzetes kísérletben egy részletes (klasszikus) reakció felállításával igazoljuk. Ezzel egyidejűleg 0,2 ml steril sóoldatot adunk a lemez minden kémcsövébe vagy üregébe, majd az első kémcsőbe vagy lyukba pipettával keverve 0,2 ml vizsgált extraembrionális vagy tenyésztő folyadékot adunk. Töltsön át 0,2 ml folyadékot a következő csőbe vagy mélyedésbe. Így a vizsgálati anyagból egymást követő kétszeres hígításokat készítünk 1:2-től 1:4096-ig vagy még nagyobb arányig. A tabletta utolsó kémcsőjéből vagy üregéből 0,2 ml anyagot veszünk a fertőtlenítő oldatba. Ezután 0,2 ml 1%-os vörösvértest-szuszpenziót adunk minden kémcsőbe vagy hígítású lyukba, a kémcsövekkel ellátott állványt vagy a lemezt megrázzuk, hogy a komponensek egyenletesen elkeveredjenek, és 22-24 °C hőmérsékleten hagyjuk. A reakciót az eritrociták önagglutinációjának szabályozása kíséri. Ehhez 0,2 ml 1%-os eritrocita-szuszpenziót adunk 0,2 ml sóoldathoz (gyakrabban 4 kémcsövet vagy lemezlyukat használnak), összekeverjük és a megadott hőmérsékleten hagyjuk. Az eritrociták agglutinációja ebben az esetben nem lehet. Az RGA eredményeit 30 perc elteltével a keresztezési pontrendszer szerint értékelik:
Négy kereszt (++++) - az agglutinált eritrociták a kút aljára telepednek le fordított esernyő formájában, szaggatott szaggatott élekkel.
Három kereszt (+++) - az agglutinált vörösvértestek esernyője kisebb, és a közepén egy nem agglutinált eritrociták korongja található.
Két kereszt (++) - az esernyő jól körülhatárolható, de a nem agglutinált vörösvértestek központi korongja megnagyobbodott és jól megkülönböztethető, a lemez megdöntésekor nem folyik.
Egy kereszt (+) - az eritrociták enyhe agglutinációja a lemez szélei mentén, amely a lemez megdöntésekor kiürül.
Negatív reakció (-) - nincs vörösvértestek agglutinációja.
A vírus vagy egy hemagglutinációs egység (1 HAU) hemagglutinációs titeréhez a vírus legmagasabb hígítását veszik, amelynél legalább két keresztezéssel vörösvértest-agglutináció figyelhető meg. Például: egyértelműen kifejezett hemagglutinációt kaptunk 1:512 hígításnál, azaz 0,2 ml izolált vírus 1:512 hígításban 1 HAU-t tartalmaz. Elfogadható, hogy az RGA-t (valamint az RZGA-t) tablettákban állítsák be biokémiai vizsgálatokhoz, az összes komponens 0,05 ml-es térfogatban történő bevezetésével.
A hemagglutináció késleltetési (gátlási) reakciója (RHGA) egy szerológiai módszer a hemagglutináló aktivitással rendelkező újonnan izolált vírustörzsek azonosítására és a referenciatörzsekkel való antigén kapcsolatuk meghatározására. De gyakrabban használják retrospektív diagnózisra, a fertőzés utáni és a vakcinázás utáni immunitás intenzitásának megállapítására a specifikus antitestek mennyiségi meghatározásával a csirkék és felnőtt madarak vérszérumában. Az RGA leállításának módja az izolált EDS-76 vírus azonosítása céljából és a betegség szerológiai diagnózisa kissé eltér egymástól. A kórokozó azonosítására ismert pozitív szérumot használnak, amely ismert titerben tartalmazza az ECC-76 vírus antihemagglutininjét. A specifikus antitestek azonosítását célzó diagnosztikai vizsgálatok során az ECC-76 vírus standard inaktivált antigénjét használják.
Az izolált vírus azonosításához először meg kell határozni hemagglutináló aktivitásának titerét az RHA-ban, kiválasztjuk a vírus 4 HAU-nak megfelelő munkahígítást (a módszert alább adjuk meg), majd 0,2 ml víruspozitív ECC-t adunk hozzá. 4 HAU vírus 0,2 ml sóoldatban 76 szérum olyan hígításban, amely nyilvánvalóan semlegesíti a bevitt vírus mennyiségét. A vírus szérummal való érintkezése után 30 percig. szobahőmérsékleten 0,4 ml 1%-os eritrocita-szuszpenziót adunk hozzá, és 30 perc elteltével az eredményt figyelembe vesszük. A hemagglutinációs késleltetés a vírus és a szérum típusának homológiáját jelzi, pl. megerősíti, hogy az EDS-76 vírust izolálták. A hemagglutinációs késleltetés hiánya azt jelzi, hogy olyan hemagglutináló kórokozót izoláltak, amely nem az ECC-76 vírus.
Az EDS-76 szerológiai diagnózisához szükséges hemagglutinációs késleltetési reakció két szakaszban történik.
Az első szakaszban az ECC-76 vírus standard inaktivált antigénjét hemagglutinációs aktivitással titrálják az RHA-ban, elkészítik és szabályozzák a munkahígítását. A második szakaszban maga az RZGA kerül.
Az RZGA-ban az antigén munkahígítását használjuk, ami 4 HAU-nak felel meg. Az antigén munkahígításának elkészítéséhez az ECC-76 vírus standard inaktivált antigénjét titráljuk az RGA-ban. Ennek eredményeként az antigén aktivitását 1 GAU-ra állítják be, majd 4 GAU-s munkahígítást készítenek. Ehhez egy ismert hemagglutináló aktivitású antigént annyiszor hígítanak fel steril sóoldattal, ahányszor a hemagglutináló titerének megfelelő 4 számmal elosztják. Példánkban tehát az antigén hemagglutináló titere (1 HAU) az RHA-ban meghatározott 1:512. Az antigén munkahígításának eléréséhez 128-szorosra kell hígítani (512:4 = 128). Ezért az RGA 1:512 aktivitású antigén munkahígításának elkészítéséhez, amely 0,2 ml-ben 4 HAU-t tartalmaz, 1 ml eredeti antigént kell venni, és 127 ml sóoldatot kell hozzáadni.
A fő reakció beállítása előtt ellenőrizni kell az antigén kiválasztott munkadózisának helyességét. Ehhez 0,2 ml steril sóoldatot adunk egy plexi lemez öt lyukába, a másodiktól kezdve. Ezután a munkahígításban lévő antigénből 0,2 ml-t adunk az első és a második mérőhelyhez. A második üregből pipettával 0,2 ml folyadékot veszünk, és átviszünk a harmadikba stb., így az antigén kétszeres hígítását készítjük el. Ezt követően 0,2 ml 1%-os csirke eritrocita szuszpenziót adunk minden egyes lyukba, a lemezt finoman rázva összekeverjük, és szobahőmérsékleten 30 percig állni hagyjuk. Nál nél jó választás az antigén munkadózisa (4 HAU) 1, 2 és 3 üregben, amelyek 4, 2 és 1 HAU-t tartalmaznak, az eritrociták intenzív agglutinációját figyelni kell. A negyedik és ötödik lyukban az agglutináció nem lehet több, mint egy kereszt. Ha a kontroll eredményei eltérnek, szükséges a standard antigén újratitrálása és a munkahígítás újbóli elkészítése.
A hemagglutinációs késleltetési reakciót plexi lemezekbe vagy kémcsövekbe helyezzük. Ezzel egyidejűleg 0,2 ml sóoldatot adunk a lemez vagy kémcső minden egyes üregébe, majd 0,2 ml tesztszérumot adunk minden sor első üregébe (kémcsőbe), és kétszeres sorozathígításokat 1 :2-től 1:4096-ig pipettával készítjük el. Ezt követően a munkahígításban lévő antigén 0,2 ml-ét minden lyukba (csőbe) adjuk, a lemezeket (állványokat) addig rázzuk, amíg a komponensek egyenletesen el nem keverednek, és 30 percig szobahőmérsékleten hagyjuk, hogy az antigén érintkezzen a szérum antitestekkel. A meghatározott idő elteltével a lemez (kémcső) minden egyes üregébe 0,4 ml 1%-os csirke eritrocita szuszpenziót adunk, a komponenseket összekeverjük és 30 percig szobahőmérsékleten hagyjuk. Ha a vizsgált vérszérumok az ECC-76 vírusra specifikus antihemagglutinint tartalmaznak, akkor az RHA pozitívnak minősül, és az antitesttitertől függően bizonyos üregekben a hemagglutináció hiányában nyilvánul meg. A szérumtiter a legmagasabb hígítás, amelynél nincs vörösvértest-agglutináció. Az RZGA beállítását vezérlőelemeknek kell kísérniük:
0,2 ml csirke eritrocita 1% -os szuszpenziója + 0,2 ml sóoldat (nem szabad az eritrociták agglutinációját);
0,2 ml EDS-76 vírusra pozitív specifikus vérszérum + 0,2 ml antigén munkahígításban + 0,4 ml 1%-os csirke eritrocita szuszpenzió (hemagglutináció nem lehet);
0,2 ml normál negatív csirkék EDS-76 vírus vérszéruma + 0,2 ml antigén munkahígításban + 0,4 ml 1%-os csirkék eritrocita szuszpenziója (egyértelműen kifejezett hemagglutinációnak kell lennie);
0,2 ml vizsgált vérszérum (tojás kivonat) + 0,2 ml 10%-os csirke eritrocita szuszpenzió (hemagglutináció nem lehet).
Az RZGA elszámolását a kontrollok elszámolásával kell kezdeni. Csak a kontroll reakció kielégítő eredményeivel lehet figyelembe venni a vizsgált szérummal végzett reakció eredményeit. Az ECC-76 vírus antihemagglutininek diagnosztikai pozitív titere 1:16 és magasabb. A tojástermelés csökkenése után 2-3 héttel vett 15-10 vérszérum- vagy tojássárgája-kivonat-minta negatív eredménye az RZGA-ban lehetővé teszi számunkra, hogy kizárjuk az EDS-76-ot, mint a csökkent tojástermelékenység egyik vezető okát.
Az EDS-76 diagnosztikai vizsgálatai akkor a legmegbízhatóbbak, ha a betegség kezdete előtt (tojástermelés csökkenése), a tojástermelés helyreállítása alatt és után csirkékből nyert páros vérszérumot vizsgálunk, körülbelül 160 éves korban tojótyúkoktól vettük. 180, 200-140 nap, 300 napos kor után és az életkor miatti rétegek működésének végén.
Az immunfluoreszcens reakciót (direkt fluoreszcens antitest módszer, MFA) használják madársejtek vagy embriók tenyészetéből izolált vírus azonosítására. A készítmények ECC-76 vírusra specifikus FITC-jelölt szérummal (antitestek fluoreszcein-izocianáttal konjugátuma) történő festése és MFA-ban történő vizsgálata az általánosan elfogadott módszer szerint történik.
Amikor a vírust fertőzött fejlődő madárembriókból izolálják, a chorioallantois membránból származó készítményeket használják a kórokozó azonosítására az MFA-ban. Az ECC-76 vírusantigén specifikus szemcsés intranukleáris zárványainak kimutatása a fertőzött sejtekben az izolált kórokozó azonosítását jelzi.
Az enzim immunoassay (ELISA) módszere egy antigén-antitest komplex kimutatásán alapul specifikus antitestek és fajellenes peroxidáz konjugátum kölcsönhatása eredményeként, amely képes a szubsztrát lebomlását előidézni, és színes termék képződését okozni. az enzimatikus reakció. A reakció beállításához a "Kit az EDS-76 diagnosztizálására enzim immunoassay segítségével" (VNIVIP) szolgál, amely immunspecifikus és nem specifikus komponensekből áll. A csirkék vizsgálati szérumából vett minták vizsgálata kétféleképpen történhet: a szérum eldörzsölése nélkül és eldörzsöléssel. Ha a vizsgált vérszérum mintáinak vizsgálatát eldörzsölés nélkül végzik el, akkor ez lehetővé teszi az ELISA-reakció eredményeinek kvalitatív értékelését az elv szerint - pozitív reakció (specifikus antitestek az EDS-76 vírussal szemben). észleltek) vagy negatív reakció (nincs specifikus antitest). Ha a vizsgálatot eldörzsöléssel végezték, akkor a szérumtitert veszik a maximális hígításnak, amelynél még mindig vannak különbségek a vizsgált szérum mintáit tartalmazó üregek tartalmának színében a negatív kontroll üregeihez képest ( negatív szérum). A betegség kezdete (tojástermelés csökkenése) előtt csirkékből nyert páros vérszérumokat célszerű megvizsgálni a betegség időszakában, illetve a tojástermelés helyreállítása után.
Az EDS-76 elektronmikroszkópos diagnosztizálása egy vírust tartalmazó anyag vizsgálatával történik az általánosan elfogadott negatív festési módszerrel, amely az anyagban 10 6 /ml víruskoncentráció mellett lehetővé teszi az elektronmikroszkópos megtekintéshez szükséges preparátumok elkészítését. 30 percen belül. Alacsonyabb vírustartalom mellett az általánosan elfogadott módszerek valamelyikével koncentrálják, vagy az immunelektronmikroszkópos módszert alkalmazzák az EDS-76 vírussal homológ csirkevérszérummal, amely a feldolgozási idő enyhe növelésével. lehetővé teszi a vírus egyidejű szerológiai azonosítását.
Bioassay 5 madár 120-130 napos korában. A vizsgálati anyagot intranazálisan vagy intramuszkulárisan adjuk be 0,3-0,5 ml térfogatban mind az öt madárnak. Ugyanennyi csirke marad a fertőzetlen kontrollcsoportban. A megfigyelési időszak 30 nap. Pozitív tesztnek tekintik a fertőzött csirkék vérszérumában az ECC-76 vírussal szembeni antihemagglutinin jelenlétét 1:16 vagy annál magasabb titerben, valamint a tojástermelékenységben és a lerakott tojás minőségében mutatkozó kifejezett eltéréseket. A fertőzött madár boncolása a szaporodási traktus szerveinek fejlődési rendellenességeit tárja fel. Ha szükséges, végezzen szövettani és elektronmikroszkópos vizsgálatokat.
Kezelés és megelőzés. Az EDS-76 megelőzése a zoohigiéniai követelmények, az általános állat-egészségügyi szabályok, a madarak tartási és takarmányozási feltételeinek szigorú betartásán, valamint az oltáson alapuló speciális betegségmegelőzésen alapul.
A szülőállomány kiegészítése az EDS-76 szerint biztonságos baromfitelepről, vagy olyan utódállat terhére szükséges, ahol speciális betegségmegelőzést végeznek. Különböző korú madarakat és különböző keresztezésű csirkéket nem kívánatos ugyanabban a helyiségben tartani. Tekintettel a csirkék kacsákból és libákból származó EDS-76 vírussal való megfertőződésének lehetőségére, a kacsa- és libafarmokat szigorúan el kell különíteni a csirkék által lakott helyiségektől, és a köztük lévő gazdasági kapcsolatokat minimálisra kell csökkenteni.
Ha fennáll az ECD-76 szövődményeinek lehetősége feltételesen patogén mikroflórával, akkor tanácsos a madárnak antibiotikumot és nitrofurán készítményeket adni 5-7 napig. Beteg és gyanús madarak esetében az A-, D3-, E-, B-vitaminok adagját emelik, a takarmány kalcium mennyiségét 2-3-szorosára növelik, és akár 0,3%-os nátrium-hidrogén-karbonátot (szódabikarbónát) adnak hozzá. a takarmány.
Az EDS-76 specifikus profilaxisára inaktivált vakcinát alkalmaznak, amelyet intramuszkulárisan vagy szubkután adnak be a madaraknak 90-120 napos korban, lehetőleg legkésőbb 30 nappal a tojásrakás megkezdése előtt. Az immunitás 14 nap után alakul ki, maximumát a 30. napon éri el. Létezik 1-, 2-, 3-, 4- és több vegyértékű vakcina, különféle kombinációkban, amelyek lehetővé teszik a madarak egyidejű vakcinázását EDS-76, Newcastle-kór, fertőző bronchitis, Gumboro-kór, "nagy fej" szindróma - pneumovírus fertőzés, reovírus fertőzés, légúti mikoplazmózis, a baromfiipar járványügyi helyzetétől függően alkalmazzák. 21-28 nap elteltével meg kell határozni az oltás utáni immunitást, és szükség esetén sürgősen újra kell vakcinázni a madarat.
Tojáscsepp szindróma-76 (ESD-76) - Tojáscsepp szindróma-76 (EDS-76) - csirkék vírusos betegsége, amelyet a tojástermelékenység csökkenése, a tojás alakjának megváltozása, a héj minősége és pigmentációja jellemez. és a fehérje szerkezetének megsértése.
Történelmi hivatkozás, eloszlás, gazdasági károk. 1975-ben számos nyugat-európai országban (Hollandiában, Angliában, Észak-Írországban, majd valamivel később Olaszországban, Franciaországban, Svédországban, Dániában, Magyarországon és más országokban) új csirkebetegség jelent meg, amelyet a tojástermelékenység csökkenése jellemez. héj nélküli vagy vékonyított depigmentált héjú tojások lerakása. A betegség különösen gyorsan terjedt azokon a területeken, ahol magas koncentráció baromfitenyésztés.
A betegségben rejlő jellegzetes sajátosságok miatt az irodalomban "tojáscsepp szindróma-76" - Tojáscsepp szindróma-76 (EDS-76) néven szerepel. Először J. Van Ecke és munkatársai írták le, 1976.
Gyorsan elterjedt, Japánban, Indiában, Ausztráliában, Nigériában, Brazíliában, Mexikóban, a Szovjetunióban és Tajvanon regisztrálták.
Az EDS-76 előfordulásának hipotézisét számos tudós javasolta, akik úgy vélik, hogy a betegség kórokozója egy kacsavírus, amely nem okoz patológiát a természetes "gazdaszervezetekben". A csirkeállományokat a Rispens törzsből származó Marek-betegség elleni vakcina alkalmazása eredményeként fertőzték meg a vírussal, amelyet adenovírussal szennyezett kacsa fibroblaszt sejttenyészet felhasználásával készítettek. A vírus gyorsan alkalmazkodott új "gazdájához", és megmutatta patogén tulajdonságait. A szerzők azzal magyarázzák az EDS-76 kórokozó kacsákból való eredetét, hogy a betegség először Hollandiában, az intenzív kacsatenyésztés országában jelent meg.
A statisztikák szerint 1993-ban Oroszországban először diagnosztizálták az EDS-76-ot csirkékben, és terápiás és megelőző intézkedéseket hajtottak végre a következő régiókban: Leningrád, Moszkva, Szverdlovszk, Cseljabinszk, Omszk - 2 140 774 csirkepopuláción. A betegek legnagyobb százaléka csirkék a leningrádi régióban (67,1%), a cseljabinszki régióban (15,9%), az omszki régióban (12,6%) voltak.
1994-ben az SSYA-76 további terjesztésére került sor - Moszkva, Leningrád, Kalinin, Vlagyimir, Vologda, Tula, Jaroszlavl, Szverdlovszk, Cseljabinszk, Nyizsnyij Novgorod, Perm, Omszk, Uljanovszk, Orenburg, Szmolenszk, Orel, Murmanszk régiókban, köztük számos autonóm köztársaságban mindössze 18 452 923 csirkét oltottak be. A betegek és a vírushordozók legnagyobb százaléka a Moszkva (21,1%), Leningrád (17,1%), Omszk (15,8%), Uljanovszk (10%) és Nyizsnyij Novgorod (8,75%) régióban volt.
1995-ben a betegséget a fenti régiókban regisztrálták, valamint a Brjanszki, Pszkov, Arhangelszk, Rjazan, Kaluga, Volgograd, Kurszk, Belgorod, Novoszibirszk régiókban, Krasznojarszk Területben stb. számos autonóm köztársaságban 52 270 669 csirkét vakcináztak. Ezek közül a legnagyobb arányban a Krasznojarszk Terület (15,5%), Moszkva (10,9%), Perm (6,2%), Uljanovszk (4,2%), Nyizsnyij Novgorod (3,6%), Novoszibirszk (3%) területen volt.
A tojáscsepp szindróma-76 jelentős gazdasági károkat okoz. Külföldi kutatók szerint a kár abból áll, hogy tojótyúkonként átlagosan 15 tojás hiányzik (átlagosan legfeljebb 50 tojást selejteznek ki a tenyészrétegekből), a héj nélküli tojások legfeljebb 15%-át, a tojótyúkok legfeljebb 12%-át tojják. vérgyűrű, a csirkék keltethetőségének 7%-os csökkenése és alacsony életképessége, ami a csirkék akár 10%-os mortalitását is növeli a két hetes korig terjedő időszakban. Az EDS-76 okozta károkat súlyosbítja a korlátozó intézkedések költségei, különösen a tenyésztési gazdaságokkal kapcsolatosak.
Kórokozó. A betegséget az Adenoviridae családba tartozó DNS-tartalmú vírus okozza.
Az ECC-76 vírust először 1976-ban izolálta J. Me. Ferran Észak-Írországban. Jelenleg a kórokozónak ez a törzse „127” szám alatt ismert. Angliában szinte egyidejűleg Bexandale izolált egy hasonló vírust a beteg csirkék leukocitáiból, az úgynevezett "BC-14". Később Magyarországon izolálták az EDS-76 vírust, és a „BB/78” nevet kapta.
A fertőzés kórokozója egy kacsa adenovírus, amely a madár adenovírusok ismert szerotípusaitól eltérően képes agglutinálni a csirkék, kacsák, libák, pulykák vörösvértesteit és a mexikói EDS-76 vírust (HO-1 törzs) képes agglutinálni az emberi vörösvértesteket, pávákat és galambokat.
Morfológiailag a virion egy körülbelül 80 nm átmérőjű részecske, amelynek fonalas folyamatai 25 nm hosszúak és 2 nm átmérőjűek, és bot alakú megvastagodásban végződnek.
A vírus jól szaporodik csirke- és kacsaembriók májának és veséjének szövettenyészeteiben, fibroblasztok tenyészetében és fejlődő kacsaembriók allantois üregében. A sejttenyészetben történő szaporodás során a vírus citopátiás hatást (CPE) vált ki, és a madárvírusokra jellemző intranukleáris zárványokat képez. Megállapítást nyert, hogy a kacsaembriók és a belőlük készített sejttenyészetek megfelelőbb modellek a vírus számára, mint a csirkeembrió sejttenyészetek. Ezt a tulajdonságot sikeresen használják vírusizolációra, magas titerű vírustartalmú anyagok kinyerésére diagnosztikai és vakcinák gyártása során.
Az SSYA-76 kórokozó viszonylag ellenáll az éternek, a kloroformnak és a hőmérsékleti tényezők hatásának. Ha a vírust 30 percig 56 °C-ra melegítjük, fertőző aktivitás lép fel
legfeljebb 50%-kal csökkenthető. A 70%-ra melegítés 10 perc elteltével a fertőző és hemagglutináló aktivitás elvesztését okozza. -25°C-on lefagyasztva a vírus több mint 3 évig életképes marad. A vírus viszonylag stabil pH 6,0-8,0 között.
Epizootológia. A tojótyúkok minden fajtája fogékonyak a betegségre, a betegség maximális manifesztációjával a legmagasabb tojástermelés időszakában (200-240 nap), de a madár a termelési ciklus bármely szakaszában érintett lehet. A betegség gyorsan terjed a szabadban tartott baromfinál. A nagyon produktív keresztezésű tojótyúkok érzékenyebbek a betegségre, ideértve Hayssex Brownt, Loman Brownt, Isa Brownt. Úgy tartják, hogy a húsos baromfi érzékenyebb a betegségre, mint a tojásos baromfi. Természetes körülmények között az EDS-76 vírus széles körben elterjedt a házi- és vadkacsák között, ugyanakkor apatogén rájuk nézve, betegséget nem okoz; de ugyanakkor fertőzési forrást is jelenthetnek a csirkék számára. A betegség csekély fertőzőképessége ellenére fennáll annak a veszélye, hogy a kórokozót vadon élő madarak: galambok, verebek juttatják be a gazdaság területére.
Egyes szerzők szerint a brojlerek potenciális fertőzésforrások lehetnek. A kórokozó fő terjedési útja a transzovariális vertikális átvitel. Amikor a betegség első klinikai tünetei megjelennek, a kórokozó vízszintes terjedése figyelhető meg.
A tojások különösen intenzíven fertőződnek a virémia időszakában, amely időben egybeesik a peterakás csökkenésével. Ugyanebben az időszakban a vírus ürülékkel, garat- és légcsőnyálkával ürül. Lehetséges, hogy kakasok spermájával izolálják. A vírus más fertőzések elleni védőoltással is továbbterjedhet.
A fertőzés kísérleti szaporodásához orális intraconjunctivalis és intracloacalis adagolási módokat, valamint beteg és egészséges madarak együttes tartását alkalmazzák.
A fertőzés előfordulására hajlamosító specifikus tényezőket az életkor kivételével nem állapítottak meg. A 76-os szindróma járványtani jellemzője a vírus azon képessége, hogy nem nyilvánul meg a szervezetben, amíg a madár el nem éri a pubertást. A vírus aktiválódásának oka a peterakás megindulása okozta stressz. 40 hetes kor után a madár nem választja ki a vírust, de specifikus antitesteket tartalmaz a vér.
A fertőzött tojásból kikelt és életük első napjaiban fertőzött csirkékből felnevelt csirkékben a tojástermelés nem csökken, bár előfordulhat, hogy nem éri el a várt szintet. Az ilyen madarak a nevelés teljes ideje alatt vírushordozók maradnak. A vírus csak 22-26 hetes korban aktiválódik és szabadul fel. A tojástermelés csökkenése csak azoknál a csirkéknél figyelhető meg, amelyek szeronegatívak a teljes tenyésztési időszak alatt, vagyis minél negatívabban reagálnak szerológiailag a csirkék egy fertőzött állományban, annál kifejezettebb a tojásesés szindróma.
Patogenezis. A látens fertőzés időszakában a vírus megmarad a bélben. A virémia az adenovírus fertőzés egyik stádiuma. Úgy gondolják, hogy a vírus újraaktiválódása a peterakás kezdetén bekövetkezett hormonális profil változása miatt következik be, amelyet stressztényezőnek tekintenek. A vírus perzisztenciájának és izolálásának időtartama fordított összefüggésben van a csirkék fertőzésének korával. Minél korábban fertőződnek meg, annál hosszabbak ezek az időszakok.
A virémia időszakában a vírusrészecskék a petevezetékek nyálkahártyájának hámsejtjeiben és más szervekben találhatók. Nyilvánvalóan a vírus közvetlen hatással van a méh mirigyhámjára, ami héj nélküli vagy vékony és törékeny héjú tojások lerakását okozza.
Bizonyítékok vannak arra, hogy a vírus megzavarja a méh nyálkahártyájának mirigysejtjeinek "nátrium-kálium pumpáját". Tehát a törött héjú tojásokat tojó csirkékben a nátrium koncentrációja a méh tartalmában jelentősen megnő, a kalcium, a kálium és a magnézium pedig csökken az ép vagy fertőzöttekhez képest, de tojásrakás normál csirkehéjjal. A savasság növekedése a kalcium feloldódásához és a héjképződés megzavarásához vezethet.
Klinikai tünetek. A betegség klinikai képe kissé kifejeződik. Nincsenek tipikus tünetek. Egyes esetekben a beteg csirkéknek hasmenése, depressziója, fodros tollazata, vérszegénysége és a takarmányfelvétel csökkenése jelentkezik.
A betegség csúcsán gyengült légzés figyelhető meg, folyékony zöldes alom 1-2 hétig. A betegség későbbi szakaszaiban a fülbevalók és a fésű cianózisa figyelhető meg. A betegség legjellemzőbb tipikus tünete a tojástermelékenység csökkenése és az alsóbbrendű tojás felvétele. A tojás termelékenysége átlagosan 15-30% -kal csökken, egyes kedvezőtlen állományokban pedig akár 50% -kal. Ugyanakkor a tojástermelés csökkenése egy bizonyos ütemben halad - a teljes csökkenési időszak 6-7 hét, ebből 4-5 hét közvetlenül a csökkenésre, 2-3 hét pedig a gyógyulási időszakra esik. . Ketrectartalommal a csirkék termőképessége szinte teljesen helyreáll, vagy 1-3%-kal alacsonyabb marad a tervezettnél, padlótartalom mellett pedig általában nem éri el 7-10%-kal az eredetit.
A betegség ideje alatt nyert tojás minőségi változása abban nyilvánul meg, hogy a beteg madár 6-8 hétig hordja a tojást héj nélkül vagy csíkos durva képződményekkel a héj felszínén. Jelentősen megnő a "márvány" tojások száma, nő a harci és rovátkolt tojások aránya. A tojások kotlási tulajdonságai romlanak, miközben a tojások termékenysége, a csirkék keltethetősége és életképessége csökken.
A héj minőségének változása, az úgynevezett "zsíros tojás" leggyakrabban húsbaromfinál és barna tojótyúkoknál figyelhető meg. A fehér leghorn fajtájú, „Haysex white” csirkéknél a tojás fehérje része, nem pedig a héja, hajlamosabb a változásra. Ezekben a csirkefajtákban a fehérje vizessé, zavarossá válik. Az EDS-76 vírus a méhbolyhok mirigyszövetének megduzzadását, majd a méh nyálkahártya epitéliumának degenerációját okozza, ami a kalcium elégtelen felszívódásához és a méh váladék pH-jának csökkenése következtében acidózishoz vezet. nyálkahártya és petevezeték. Ez viszont a héjmirigy által termelt kalcium-karbonát semlegesítését okozza, ami héj nélküli, puha, vékony, depigmentált héjú tojásokat eredményez.
A tojáscsepp-szindrómás-76-os madarak mortalitása még a betegség csúcspontján is elenyésző. A kifejlett rétegek mortalitása 3-5% lehet, főként a sárgája hashártyagyulladása és a megnövekedett csípések miatt.
kóros elváltozások. Az ECD-76 fő patomorfológiai változásai a reproduktív traktus szerveiben - a petefészekben, a petevezetékben, a méhben - lokalizálódnak.
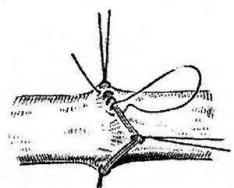
Az EDS-76 vírus által érintett, elhullott és erőszakkal megölt felnőtt madár boncolása során minden esetben kisszámú érlelő és érett tüszőt észlelnek a petefészekben. Gyakran megfigyelhető a petefészek atrófiája, gumós, deformált és sorvadt tüszők jelenléte bennük, szürkéssárga vagy zöldes színű, alvó állagú tömeggel töltve. A kötőszöveti kapszulában hiperémiás vagy vérzéses tüszők vannak. Az érintett tojótyúkok petevezetéke általában lerövidül, fala elvékonyodik, egyes esetekben hiperémia gócokkal. Figyelembe véve a folyamat lokalizációját és a betegség etiológiáját, egyes szerzők azt javasolják, hogy nevezzék "adenovírus salpingitisnek".
Sok esetben a boncoláskor a májban változást észlelnek, amely megnövekszik, ödémás, petyhüdt állagú és sárgás agyag színű. Változás van az epehólyagban, amely megnagyobbodott, vizes konzisztenciájú, világoszöld színű epével van tele. A hipertrófiás máj és az epehólyag normál működésének megsértése általános emésztési zavarokhoz vezet, ami a bél petyhüdtségében és mozdulatlanságában fejeződik ki, habzó zöldes színnel és emésztetlen élelmiszer vizes állagával töltve.
szövettani változások. Az érintett madár szerveinek és szöveteinek szövettani vizsgálata feltárja a főbb változásokat, elsősorban a szaporodási traktus szerveiben, valamint a májban.
Tehát a petefészkekben limfoid-hisztiocita és mononukleáris proliferáció, hiperémia, dystrophia és a betegség későbbi szakaszaiban nekrotikus változások figyelhetők meg az egyes tüszők epiteliális rétegében. A petevezeték fehérje részében mononukleáris limfoid infiltrációs gócok is találhatók, amelyek a nyálkahártya redőinek kötőszöveti septumában perivaszkulárisan képződnek, és annak mirigyszövetére is kiterjednek. A méhben nyálkahártya ödéma és fokális limfoid-hisztiocita proliferáció van. A májban hiperémia, perivaszkuláris proliferáció jelenléte, a hepatociták dystrophiás változásai, gyakrabban zsíros jellegűek.
Immunitás. Az ECC-76 vírus törzsei a fertőzést követő 7-14. napon specifikus antitestek képződését idézik elő a fertőzött madárban - anti-hemagglutináló, kicsapó és vírussemlegesítő.
Diagnosztika. Az EDS-76 diagnózisát járványtani, klinikai, patoanatómiai adatok és laboratóriumi eredmények alapján állítják fel.
A baromfitojás termelékenység grafikonjainak értékelése fontos az előzetes diagnózis felállításához. Mivel az EDS-76 vírus a 200-240 napos madarakban a tojástermelés 15-30%-os csökkenését idézi elő, a tojótyúkok máskor, különösen 300 napos kor utáni produktivitásának csökkenése más tényezők okozója. A diagnózis felállításában nem kis jelentőségű a tojás minőségének értékelése - alakja, színe és héj minősége, fehérje minősége.
A laboratóriumi módszerek döntőek az EDS-76 diagnózisában.
1. Specifikus antitestek kimutatása beteg és gyanús tojótyúkok vérszérumában hemagglutinációs késleltetési teszttel (HRGA) és enzim-linked immunoszorbens assay (ELISA) alkalmazásával.
A vizsgálathoz páros vérszérumot vesznek, 160-180 napos kortól, mindenesetre a tojástermelés visszaesésének kezdetétől. Ha az antihemagglutináló antitesteket 1:4-es és afeletti titerekben mutatják ki, az ECL-76 kórokozó perzisztenciájára vonatkozó következtetést vonunk le.
2. A vírus izolálása a kóros anyagból, majd az RZGA-ban történő azonosítása.
A vírus kóros anyagból történő izolálása a legmegfelelőbb 10-12 napos kacsaembriók fejlődésében, vagy fibroblaszt sejttenyészetben, kacsaembriók májában, vesében.
3. Differenciáldiagnózis. Az EDS-76 differenciáldiagnózisában a tojástermelés CELO-vírusok, fertőző hörghurut, Newcastle-betegség, valamint a tartási és takarmányozási feltételek megsértése miatti csökkenése kizárt.
specifikus profilaxis. Az EDS-76 megelőzésére ásványi és olajadjuváns alapú, monovalens és kapcsolódó oltóanyagokat fejlesztettek ki és vezettek be a baromfitenyésztés gyakorlatába. A madarakat 100-110 napos korukban egyszer, intramuszkulárisan vakcinázzák.
A fertőző bronchitis, az IBD, a Newcastle-betegség és az EDS-76 elleni kapcsolódó inaktivált vakcinákat mára széles körben használják.
Annak a személynek, aki a telephelyén tojás- és húscsirkéket tenyészt, nemcsak a tenyésztési és karbantartási szabályokat kell megtanulnia, hanem ismernie kell azokat a betegségeket is, amelyek tollas kedvenceit érinthetik. És nem csak tudni kell róluk, hanem időben és helyesen reagálni, hogy ne hagyja ki a madár életére, valamint az emberi egészségre veszélyes körülményeket. Ez a cikk a tojáscsepp szindróma-76 nevű gyakori betegséget tárgyalja.
Vannak betegségek baromfi fajról a másikra terjednek, kifejezett tünetek nélkül, amíg nem találnak egy áldozatot, aki érzékeny a betegség kórokozójára.
Tudtad? A csirkéket először mintegy háromezer évvel ezelőtt háziasították a mai Etiópia területén.
A tojáscsepp-szindrómát (ESD-76) először 1976-ban fedezték fel és írták le Hollandiában. Általánosan elfogadott, hogy a kacsák hordozzák a vírust: házi és vadon élő kacsák, ők maguk azonban nem fogékonyak a betegségre.
Az a tény, hogy a meghatározott év előtt csirkevérből vett szérummintákban nem találtak a kórokozó ellen antitesteket, annak bizonyítéka, hogy ez a betegség ebben az időszakban keletkezett.
Ezt követően az eredeti-127-es törzzsel azonos vírustörzseket izoláltak különböző progresszív országokban: Angliában, Franciaországban, Olaszországban, Japánban és Magyarországon. Ez azt jelenti, hogy a felfedezett betegség elkezdett terjedni az egész világon. 
Az ESS-76, vagyis az adenovírus betegség (Egg drop Syndrome-76) jellemzője, hogy a tojótyúkoknál a reproduktív rendszer károsodása miatt csökken a tojástermelékenység, a tojások alakja megváltozik, minőségük romlik, a héj töredékesen depigmentált és meglágyul vagy teljesen hiányzik, a fehérje szerkezete megbomlik.
Ennek a patológiának a kórokozója egy DNS-tartalmú adenovírus (Adenoviridae), innen ered a betegség másik neve. Ez a mikroorganizmus nem tartozik a madár-adenovírusok ismert típusai közé, és az említettektől eltérően képes sok madár vörösvértestének összeragasztására (agglutinációjára), beleértve a házimadarakat is.
Tudtad? A csirke nem fog feküdni a sötétben, még akkor sem, ha ennek eljött az ideje. Megvárja, hogy eljöjjön a nap, vagy hogy felgyulladjanak a lámpák.

Miután a csirke elszenvedte ezt a betegséget, antitestekre tesz szert, amelyeket a tojáson keresztül továbbíthat az utódoknak.
A mikroorganizmus érzékeny a formaldehidre, de nem pusztítható el:
- éter;
- kloroform;
- tripszin;
- 2%-os fenolos oldat;
- 50%-os alkoholos oldat.
50 fokon 3 órán át aktív, 56 fokon - egy órát, 80 fokon - fél órát. Ismeretes, hogy a kórokozó a petevezeték hámjának sejtjeiben felszaporodik, és ezzel egyidejűleg a normál minőségű tojáshéj képződése is megzavarodik.
Tudtad?Egy egynapos csaj olyan reflexekkel és készségekkel rendelkezik, amelyek megegyeznek egy hároméves embergyerekével.

Beteg madárnál a gyógyulás után a következők figyelhetők meg:
- a petevezetékek ödémája és a bennük lévő atrófiás folyamatok - lerövidülés és elvékonyodás;
- bizonyos esetekben - ciszták;
- változások a májban: méretnövekedés, sárgás, petyhüdt szerkezet;
- az epehólyag megnagyobbodása és folyadékkal való feltöltése.
A betegség okai
Bármilyen fajtájú és életkorú csirke megbetegedhet, a produktív kortól kezdve, azonban a vírus megnyilvánulásának "kedvenc" életkora a csirke termelékenységének csúcsa: 25-35 hét. Nagyobb fogékonyságot mutatnak rá a tenyésztenyésztési csirkék, valamint a hústípusba tartozó tojótyúkok.
Minél világosabbak a betegség megnyilvánulásai, annál nagyobb termőképesség várható az egyedtől fajtajellemzőinek megfelelően.  A transzovariálisan (fertőzött tojótyúk által lerakott tojáson keresztül) terjedő adenovírus tünetmentesen maradhat a fiatal madár szervezetében egészen addig az időpontig, amíg szervezete stresszt tapasztal, például a peterakás megindulásáig. A számára megfelelő pillanatban aktiválódik, csökkentve a csirke tojástermelését. Ezt az átviteli módot vertikálisnak nevezik.
A transzovariálisan (fertőzött tojótyúk által lerakott tojáson keresztül) terjedő adenovírus tünetmentesen maradhat a fiatal madár szervezetében egészen addig az időpontig, amíg szervezete stresszt tapasztal, például a peterakás megindulásáig. A számára megfelelő pillanatban aktiválódik, csökkentve a csirke tojástermelését. Ezt az átviteli módot vertikálisnak nevezik.
Figyelemre méltó, hogy a fertőzött tojásból kikelt vagy az EDS-76 kórokozóval fertőzött csirke élete első napjaiban nem mutatja a szindróma egyértelmű megnyilvánulását a termelékenység csúcsán, azonban nem kell tőle magas tojástermelést várni. .
A horizontális fertőzés lehetősége is fennáll:
- érintkezés - emberek ruházatán és cipőin keresztül, szállításon, háztartási cikkeken és gondozáson keresztül;
- szexuális - egy kakas spermáján keresztül;
- széklet-orális - ürüléken és fertőzött egyének orrüregéből és szájüregéből való váladékon keresztül;
- a madarak egyéb betegségek elleni vakcinázásával.

Az SSJ-76 kórokozó hordozói fertőzöttek, valamint beteg csirkék, kacsák és libák, házi és vadon élő állatok, valamint egyéb vízimadarak. A fertőzött ürüléken keresztül a vadon élő madarak nagy távolságokra hordozhatják a betegséget.
Fontos! Ha a madarakat zsúfolt, szoros érintkezésben tartják, a vírus terjedése nagymértékben felgyorsul, és 1-14 napon belül a teljes állomány fertőzése bekövetkezhet. Ezzel szemben az egymástól válaszfalakkal elkülönített tojótyúkok hosszú ideig egészségesek maradhatnak, még a fertőzött egyed közelében is.
Gazdasági kár
Az EDS-76 jelentős gazdasági károkat okoz mind a magángazdaságoknak, mind a nagy ipari vállalkozásoknak. A betegség során egy tojótyúk selejtezése 10-30 tojás, tenyészmadaraknál eléri az 50-et. Ez 17-25%-os veszteséget jelent.  Egy egyed termőképességének helyreállítása ketrecben tartva 4-6 hétig tart. A földön tartott, más egyedekkel és azok biológiai anyagával érintkező tyúkoknál előfordulhat, hogy a tojástermelés 6-12%-kal soha nem áll vissza az eredeti szintre.
Egy egyed termőképességének helyreállítása ketrecben tartva 4-6 hétig tart. A földön tartott, más egyedekkel és azok biológiai anyagával érintkező tyúkoknál előfordulhat, hogy a tojástermelés 6-12%-kal soha nem áll vissza az eredeti szintre.
Ami a fertőzött egyedek által lerakott keltetőtojásokat illeti, sok közülük egyszerűen alkalmatlan fiatal állatok tenyésztésére a túl törékeny héj miatt. Amellett, hogy nagy százalékban már a kezdeti szakaszban selejteznek, a fiókák keltethetősége is csökken. A kikelés utáni első napok túlélési szintjét is csökkentették.
Bár korunkban sokkal több információ áll rendelkezésre erről a betegségről, és 1976-hoz képest elegendő tapasztalat halmozódott fel a harcban, néhány kérdés még mindig ellentmondásos, és nincs egyértelmű válasz.
Fontos! A szindróma elterjedt azokban az országokban, ahol magasan fejlett ipari baromfitechnológiát alkalmaznak, és a legnagyobb károkat a tenyésztelepeken okozza.

Tünetek
A fertőzött egyedben a produktív kor kezdete előtt a kórokozó a bélben tartózkodik, és semmilyen módon nem nyilvánul meg. Amikor eljön az ideje, és a csirke hormonális háttere megváltozik a tojástermelés biztosítására, a vírus aktiválódik, és megkezdődik a virémia stádiuma, vagyis a vírus keringése a szervezetben a véráramon keresztül.
A petevezeték nyálkahártyájának hámrétegét elérve a vírus hozzájárul az ásványi anyagok egyensúlyának felbomlásához: nátrium, kálium, magnézium, kalcium és mások, aminek következtében a csirke túl vékony, deformált vagy akár teljesen hiányzó héjú tojásokat rak. .
Tudtad? A kakas a csirkeállományban a reproduktív szerepe mellett számos fontos társadalmi és adminisztratív funkciót is ellát: a napi rutin irányítása, a konfliktusok megelőzése, a veszélyek elleni védekezés, még akkor is, ha az ellenség erőben és méretben nyilvánvalóan meghaladja őt.

A fertőzés súlyossága ellenére a csirkéken ritkán mutatkoznak a betegség jelei.
Alkalmanként, gyakrabban kisebb formában, előfordulhatnak:
- általános mérgezés jelei - gyengeség, fáradtság és mások;
- étvágytalanság;
- hasmenés és zöld árnyalat jelenléte az alomban;
- anémia;
- gyenge légzés az akut állapot csúcsán;
- cianotikus árnyalatú fésű és fülbevaló.
A fő jel és tünet a termelékenység éles csökkenése, a vékony, deformált, nagyon rossz minőségű tojások lerakása. Az ilyen termékben lévő fehérje vizes és zavaros. Az ezekből a tojásokból kikelt csibék életképessége alacsony, és életük első napjaiban nagy számban pusztulnak el.  A tünetek a csirke fajtájától függően változhatnak:
A tünetek a csirke fajtájától függően változhatnak:
- A „zsírtojás” és a csökkent héjminőség gyakrabban fordul elő barna keresztezetteknél és brojlercsirkéknél;
- fehérje változás. Vékonyodása, zavarossága inkább a fehér keresztekre jellemző.
Fontos! Az eset nem jellemző tünete ennek a betegségnek, szintje ritkán haladja meg az 5%-ot. Az ok elsősorban a sárgája hashártyagyulladása.
Diagnosztika
Az előzetes diagnózis felállítása és a nyomon követési nyilvántartás vezetése érdekében a tojástermelékenység alakulását ábrázoló grafikonokat kell kidolgozni, figyelembe véve, hogy az adenovírus miatt a tojótyúkoknál 200-240 tojástermelés csökkenés következik be. napos korban.
Abban az esetben, ha egy 300 naposnál idősebb egyén termelékenysége csökken, az ok valószínűleg más tényező.  Mindenesetre a "tojáscsepp-szindróma-76" diagnózisa előtt ki kell zárni a következőket:
Mindenesetre a "tojáscsepp-szindróma-76" diagnózisa előtt ki kell zárni a következőket:
- fertőző etiológiájú hörghurut;
- mérgezés különböző anyagokkal;
- nem megfelelő étrend;
- egyéb tényezők, amelyek a tojástermelékenység csökkenését okozhatják.
Hogyan és hol kell jelentkezni
Ha egy ipari vállalkozásnál észlelik a vírust, a gazdaság a hátrányos helyzetűek kategóriájába kerül, és megfelelő korlátozásokat vezetnek be: mechanikus tisztítási és fertőtlenítési intézkedések, vakcinázás, selejtezés és hasonlók.
Az EDS-76-gyanús csirke felfedezése egy privát csirkeólban alkalom arra, hogy meghívjunk egy állatorvost, aki megvizsgál, beolt, és ajánlásokat ad. 
Milyen vizsgálatokat fognak végezni
Az "adenovírus fertőzés" diagnózisa a következő vizsgálatok alapján történik:
- járványtani;
- klinikai;
- kóros;
- laboratórium.
A laboratóriumi elemzéshez vizsgálja meg:
- petevezeték;
- petefészkek tüszőkkel;
- végbél és annak tartalma;
- vér;
- tamponok a nasopharynxből és a kloákából.
Célszerű a betegség első napjaiban (3-5 nap) kutatást végezni, és olyan madárból származó anyagot használni, amely legfeljebb 2 órával ezelőtt elhullott vagy levágott. 
Izoláláshoz és szérumának vizsgálatához az alábbi csoportok egyedeitől tanácsos vért venni (15-20 minta mindegyikből):
- 1-200 napos egyedek;
- 160-180 napos egyedek;
- 220 napos egyének;
- 300 napos egyének;
- szolgálatból kivont idős személyek;
- betegség jeleit mutató példányok.
Tudtad? A csirkéknek megvan a saját „nyelvük”, amely mintegy 30 féle jelet képes hang segítségével továbbítani a többi egyednek, sőt létezik egy „anyanyelv”, amelyet az anya tyúk kommunikál utódjával. Sőt, néhány nappal ez előtt az esemény előtt egy fióka, amelyik még nem kelt ki, kommunikál a tyúkkal a héjon keresztül, és ehhez akár tíz különböző jelet is felhasznál.

Ami a tojást illeti, kívánatos a nem szabványos minták tanulmányozása, amelyek megsértik a héj szerkezetét és / vagy a tartalmat.
Hogyan kell kezelni
Sok más vírusos betegséghez hasonlóan nincs specifikus kezelés. Javasolt az étrend hasznosságára, esszenciális aminosavakkal, vitaminokkal és ásványi anyagokkal való telítettségére összpontosítani. Az antitestek termelése a betegség 5-7. napján kezdődik és 2-3 hétig tart, majd az egyén életre szóló immunitást kap.
A szükséges intézkedések közé tartozik az első megbetegedett tojótyúkok kötelező elkülönítése az állomány többi részétől, különösen, ha padlón történő elhelyezést alkalmaznak. Ebben az esetben meg kell figyelni a madár többi részét a tünetek jelei miatt.
Ha a betegség természete nem egyedi, karantén intézkedésekre van szükség. A működésképtelen madarat le kell vágni, a belőle levett biológiai anyagot elemzésre küldik a diagnózis laboratóriumi megerősítésére. 
Fontos! Nagyon fontos, hogy nyomon kövessük a betegség kezdetét, és ne kezdjük el a helyzetet: ezzel elkerülhetjük a vírusnak a csirkeállományban való terjedésével kapcsolatos számos problémát.
Ez az intézkedés hatékony lehet a virémiás fázis kihagyásában – a vírus véráramon keresztüli terjedésében az egész szervezetben. Következésképpen a kórokozó kevésbé károsítja a madarat, nem lesz jelen a testváladékban, ráadásul ez az intézkedés javítja a tojások minőségét és a madár produktivitását. 
Megelőzés és védőoltás a vírus ellen
Az olyan kellemetlen betegség megelőzésére, mint a tojáscsepp szindróma-76, vakcinázást alkalmaznak, amely megakadályozza a virémiás fázist, ami javítja a tojáshéj termelékenységét és minőségét.
A 16-20 hetes egyedeket szubkután vagy intramuszkulárisan injekciózva kell beoltani, és 2 hét elteltével a madár egy évig tartó immunitást alakít ki.
Az immunizáláshoz a következő vakcinákat használják:
- folyadék inaktivált;
- emulgeált inaktivált;
- asszociatív inaktivált.

A megelőző intézkedések az állat-egészségügyi szabályok betartásán alapulnak annak érdekében, hogy megakadályozzák a kórokozók külső környezetből való bejutását. A csirkék keltetéséhez 40 hetesnél idősebb tojótyúkoktól vett tojásokat használnak, és először meg kell győződni arról, hogy az elemzésük normális.
A madarat, amelynek vérében megtalálják a kórokozót, levágják. A vírus kimutatásának ténye jelzi a vírus jelenlétét a gazdaságban. Ebben az esetben figyelnie kell a csirkeólját, és időben meg kell tennie a szükséges intézkedéseket.
A betegség kitörésének kockázatának csökkentése a csirkeólban:
- megfelel az egészségügyi szabványoknak;
- a madarakat korcsoportok szerint elkülönítve kell tartani;
- a csirkeállományt külön kell tartani a libától és a kacsától;
- időnként takarítsa és fertőtlenítse a helyiséget, valamint a leltárt.
Tudtad? A csirkék képesek érzelmekre: együttérzés, szomorúság. Emellett megfelelő szintű intelligenciával rendelkeznek ahhoz, hogy emlékezzenek körülbelül száz másik lény megjelenésére, és a környezettel kapcsolatos tapasztalataikat és információikat is felhasználják a döntések meghozatalához.
5
alkalommal már
segített
Átalakító